|
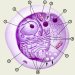
Клітка (cytus) - основна структурно-функціональна одиниця, що визначає будову, життєдіяльність, розвиток і розмноження тваринних і рослинних організмів, за винятком вірусів; елементарна жива система, здатна до обміну речовин з навколишнім середовищем і до самовідтворення. К. можуть існувати як самостійні організми (бактерії, одноклітинні водорості, гриби, найпростіші) або утворюють тканини багатоклітинних організмів. Наука про клітину - цитологія - має ряд напрямків, головними з яких є цитоморфология, цитофизиология і цитопатология.
В залежності від рівня клітинної організації розрізняють два типи клекти: прокаріоти, або доядерные, і еукаріоти, або ядерні. Прокариотические клекти типові для синьозелених водоростей і бактерій, эукариотических кліток складаються всі інші рослинні і тваринні організми.
Основними структурами эукариотической клекти є ядро, занурене в цитоплазму, мембранна система, всіх органел, а також спеціалізовані структури.
Ядро клекти складається з кариоплазмы (нуклеоплазми), одного або декількох ядерець та ядерної оболонки. Як правило, в клекти є лише одне ядро, але існують дво - та багатоядерні клітини.
Кариоплазма містить всю хромосомну ДНК клітини, яка в асоціації з основними білками - гистонами - утворює нитки хроматину - нуклеогистоны. У цих нитках гистоны формують дископодібні комплекси, навколо яких обвиваються суперспирали ДНК. В результаті виникають повторювані структури типу бус - нуклеосомы; останні, в свою чергу, складаються у вигляді спіралі, завдяки чому досягається надзвичайно компактна упаковка гігантських молекул ДНК, довжина якої в кожній хромосомі в середньому дорівнює 5 див. В залежності від рівня конденсації ДНК розрізняють еухроматин, тобто зони повної деконденсации, і гетерохроматин, де упаковка нуклеосом настільки щільна, що їх комплекси видно в світловий мікроскоп як гранули різних розмірів. У кариоплазме між нитками хроматину містяться кислі (негистоновые) білки, багато з яких беруть участь у регуляції матричної діяльності ділянок ДНК (генів), гранули рибонуклеопротеинов (РНП), різні ферменти та ін Крім того, тут виявляється система білкових ниток діаметром 2 нм, які об'єднуються у фібрили вищих порядків діаметром 20-30 нм, формуючи структурний матрикс ядра. Під оболонкою ядра матрикс згущується в щільну пластинку, з якою пов'язані ділянки хромосом і порові комплекси оболонки ядра. Уздовж хромосом один за одним розташовуються ділянки, в яких здійснюється самовідтворення (реплікація) молекул ДНК; ці ділянки називають репликонами.
В ядрі знаходяться ядерця, містять РНК; розміри ядерець (1-5 мкм) можуть варіювати в залежності від стану клітини.
Ядро оточене ядерною оболонкою, що складається з внутрішньої і зовнішньої мембран; простір між ними називається перінуклеарним. Ядерна оболонка бере активну участь в обміні речовин між ядром і цитоплазмою.
У цитоплазмі розрізняють гиалоплазму, мембранну систему, всіх органел і різноманітні включення. Гиалоплазма (цитозрль, матрикс цитоплазми) - складна биоколлоидная система, яка об'єднує всі структури клекти і служить середовищем для їх хімічної взаємодії. Важливу роль у цих процесах відіграють розчинені в цитозолі ферменти і АТФ. Крім того, гиалоплазма містить субмикроскопическую тривимірну мережу білкових волоконец трьох типів: микрофиламенты (актиновые филаменты) діаметром 4-7 нм, побудовані з сократимых білків, проміжні филаменты (микрофибриллы) діаметром близько 10 нм і мікротрубочки - негільчаті порожнисті циліндри діаметром близько 25 нм. Ряд допоміжних білків пов'язує ці волоконця в каркас клекти - цитоскелет; по периферії клекти згущення філамент формує термінальну мережу. Зміни в цитоскелете впливають на в'язкість цитоплазми; локальні зміни в'язкості разом з скороченнями микрофиламентов призводять до руху клекти. Цитоскелет також бере участь у внутрішньоклітинному транспорті речовин і, можливо, в просторовій організації ферментних систем клітини.
Покриває клітку оболонка (плазматична мембрана, плазмолемма) являє собою комплекс ліпідних і білкових молекул. Перші утворюють безперервний подвійний шар товщиною 6-10 нм (ліпідний бислой), в який занурені молекули структурних, транспортних, рецепторних та інших білків; ліпідний бислой визначає структурні особливості мембрани, а білки - більшість її функцій. На зовнішній поверхні К. виступають ланцюга вуглеводів, сполучених з білками (глікопротеїни) або з ліпідами (гликолипиды) плазмолеми; ця зовнішня зона товщиною 3-4 нм називається глікокалікс.
Спеціалізовані ділянки плазматичних мембран дотичних К. утворюють міжклітинні контакти різної будови. На вільній поверхні ряду типів К. є мікроворсинки, війки, джгутики та ін.
Подібно цитоскелету, плазматична мембрана - динамічна структура, яка регулює взаємодію між клітинами і її оточенням, включаючи інші клітини. При проходженні частинок і макромолекул всередину клітини (эндоцитоз) або при виведенні їх з клітини. (экзоцитоз) від плазмолеми безперервно відриваються одні ділянки і вбудовуються в неї інші (кругообіг мембранних компонентів клітини). Процес эндоцитоза поділяють на фагоцитоз (захоплення корпускулярних частинок) і пиноцитоз (захоплення колоїдів). Ділянку мембрани, отрывающийся в цитоплазму разом із захопленим матеріалом, утворює фагосому; надалі перетравленні вмісту фагосомы беруть участь ферменти лізосом. Захоплення частинок і макромолекул на поверхні клітини забезпечується спеціальними рецепторными білками. Після цього рецептори збираються в маленькому ділянці мембрани, під яким накопичується білок клатрін, формує тут облямівку. Занурюючись у клекти, така ділянка утворює облямовану ямку, де концентруються однорідні, відповідні рецептора макромолекули, що поглинаються майже без позаклітинної рідини (рецепторний эндоцитоз).
З плазматичної мембраною подібні за будовою внутрішні мембрани клекти - цитомембраны. Вони поділяють внутрішній простір клекти вищих організмів на відсіки - компартменты, які організовують процеси обміну і забезпечують можливість одночасного перебігу в одній клекти багатьох хімічних реакцій, часом несумісних один з одним. Ці відсіки називають мембранними органоидами.
Всіх органел (органели) цитоплазми являють собою постійні структури клітини, що виконують певні функції. До них відносяться ендоплазматичний ретикулум, комплекс Гольджи, мітохондрії, рибосоми та ін.
Ендоплазматичний ретикулум (ендоплазматична мережа) являє собою систему цистерн, канальців і вакуолей, обмежених цитомембраной. Розрізняють гранулярний (шорсткий) і агранулярный (гладкий) ендоплазматичний ретикулум; у першому переважають плоскі мішечки - цистерни, у другому - канальці. Мембрани шорсткого ретикулума з боку гиалоплазмы покриті рибосомами. Ступінь розвитку цього органоида залежить від рівня метаболічної активності і диференціювання клітини: він сильніше розвинений у клітинах, активно синтезують білки.
Комплекс Гольджи - система сплощених цистерн і пухирців, оточених мембраною; зазвичай знаходиться недалеко від ядра. У секретують клекти цей органоид розташовується на зверненої до зовнішнього середовища стороні ядра. Від структур комплексу Гольджи відокремлюються бульбашки, які, зливаючись, формують секреторні гранули.
Мітохондрії оточені подвійною оболонкою, що складається з зовнішньої і внутрішньої мембран; їх розмір 0,2-2,0 мкм, тобто вони видні у світловий мікроскоп. Внутрішня мембрана утворює складки (крісти), вдающиеся у заповнює внутрішню камеру мітохондрії матрикс; останній містить кільцеві молекули ДНК і мітохондріальні рибосоми. У рослинних клекти цим органоидам відповідають пластиды, здійснюють фотосинтез. Головна функція мітохондрій - участь в енергетичному обміні.
Лізосоми - оточені одинарною мембраною тільця розміром 0,2-0,4 мкм. В них знаходяться ферменти (гідролази), які здійснюють розщеплення макромолекул і часток як захоплених клекти ззовні, так і завершили цикл свого існування в клекти. Первинні лізосоми розвиваються в комплексі Гольджи. Зливаючись з фагосомами, вони утворюють вторинні лізосоми, в яких відбувається переварювання і засвоєння захоплених речовин. Частина лізосом (аутолизосомы, цитолизосомы) перетравлюють відмираючі структури самої клекти. Лізосоми з залишками неперетравлених речовин називають телолизосомами (залишкові тільця).
Рибосоми - немембранные всіх органел, які містяться в усіх живих клекток, це універсальний апарат синтезу білкових молекул. Вони побудовані з білків і РНК; в функціонуючому стані складаються з великої та малої субодиниць і мають розміри 252020 нм.
Микротельца - загальна назва оточених мембраною пухирців діаметром 0,1-1,5 мкм. В їх серцевині знаходяться ферменти, що каталізують різні окислювальні реакції. Наприклад, ферменти пероксисом каталізують утворення і руйнування Н2О2, що використовується в ряді метаболічних циклів.
Клітинний центр (центросома) складається з пари центриолей (диплосома), оточених тонковолокнистой зоною (центросфера). Кожна центриоль має форму циліндра розміром близько 0,3-0,51,5 мкм. Цей органоид - центр організації мікротрубочок цитоплазми; він пов'язаний з розвитком війок і джгутиків, утворенням веретена поділу і в цілому з координацією рухів клітини.
У деяких клектах (м'язових, нервових), крім названих общеклеточных органоїдів, є спеціальні всіх органел (міофібрили, синаптичні пухирці), пов'язані з виконанням К. її специфічних функцій.
Цитоплазма може містити включення - щодо непостійні компоненти, які утворюються і зникають у ході клітинного метаболізму. Вони видні у світловий мікроскоп як щільні (гранули) або рідкі (вакуолі) грудочки і краплі. Розрізняють трофічні (білки, жири, глікоген), секреторні, экскреторные і пігментні включення.
Все розмаїття клекток тваринного організму виникає шляхом їх розвитку з однієї заплідненої яйцеклітини В ході поділу функцій (спеціалізації) К. купують одні і втрачають інші структури. В результаті процесу диференціювання у людини утворюється близько 1013 клітин, які прийнято об'єднувати в 4 тканинних типи: епітеліальні, сполучнотканинні, м'язові і нервові. Розміри клекти коливаються від 4-6 мкм (малі лімфоцити людини) до декількох сантиметрів (яйцеклітини птахів); довжина відростків нервових клекток досягає 1,5 м.
Биосинтетические процеси в клітині. Генетична інформація молекул ДНК реалізується шляхом синтезу на матриці расплетенных ниток ДНК молекул РНК. Цей процес називають транскрипцією. По мірі освіти нитки РНК з'єднуються з білками в рибонуклеопротеиновые комплекси (РНК). У транскрипції бере участь дуже мала частина клітинної ДНК эухроматина. Спочатку утворюються довгі молекули так званої гетерогенної ядерної РНК; в ході дозрівання (процесингу) з неї, як правило, вирізаються та видаляються значні ділянки (интроны). Зрілу РНК, яка служить матрицею для синтезу білків у цитоплазмі, називають інформаційної РНК (иРНК). Крім неї на матрицях особливих генів синтезуються транспортні РНК (тРНК) і рибосомные (рРНК). Виходячи в цитоплазму, молекули тРНК з'єднуються в цитозолі з амінокислотами, причому кожному виду тРНК відповідає своя амінокислота, а комплекси рРНК з білками формують малі і великі субодиниці рибосом.
Синтез білка на матриці иРНК (трансляція) протікає в цитоплазмі. Білки, що йдуть в основному на потреби самої К., синтезуються на вільних рибосомах цитозолю; при синтезі мембранних або секреторних (підлягають виділенню) білків рибосоми прикріплюються до мембран ендоплазматичного ретикулума, складаючи їх гранулярний компонент. Ліпіди утворюються на мембранах агранулярного ретикулума. Синтезовані речовини надходять в порожнині ретикулума і транспортуються в комплекс Гольджи; по мірі просування вони дозрівають і в кінцевому рахунку формують секреторні пухирці, що прямують до плазматичної мембрани. Перенесення речовин між органоидами К. здійснюють оточені мембраною транспортні бульбашки.
Процеси біосинтезу йдуть з поглинанням енергії, основним постачальником якої є мітохондрії. Тут енергія, те що вивільняється в ході окислювальних реакцій, вловлюється і накопичується у вигляді молекул АТФ; у міру потреби ці молекули, розщеплюючись, звільняють необхідну енергію. Деяку роль в обміні енергії відіграє також АТФ цитозолю.
Таким чином, інформація, що надходить з ядра, визначає (детермінує) синтез макромолекул в цитоплазмі. Однак частина проектів біосинтезу надходить в ядро, здійснюючи регуляцію за типом зворотного зв'язку або безпосередньо, як, наприклад, гистоны, включаючись до складу хромосом. Вплив мікрооточення на життєдіяльність і диференціювання клітини також передається через цитоплазму.
Ядро і цитоплазма складають нерозривне ціле. Кожен з цих компонентів окремо може жити лише обмежений час і не здатна до самовідтворення, тому будь-який прояв життєдіяльності клітини - результат взаємодії її взаємопов'язаних компонентів.
Биосинтетические процеси йдуть в клекте безперервно, поки в ній зберігається оформлене ядро (період інтерфази) і практично припиняються під час поділу клітини (період мітозу). Період часу від виникнення клекти в результаті поділу материнської клітини до її власного поділу або загибелі називають клітинним циклом. Послідовні періоди циклу позначають символами G1, S, G2 і М.
У G1 (пресинтетический період) синтезуються й нагромаджуються речовини, необхідні для реплікації ДНК. У пізньому G1-періоді настає момент (точка рестрикції, R-точка), коли клітини може вийти з циклу і виконувати свої специфічні функції, не пов'язані з поділом. Такі «спочиваючі клітини називають клітинами, що знаходяться в періоді G0 циклу. У цьому стані клітини можуть перебувати невизначено довго; при цьому одні К. (наприклад, нервові) втрачають здатність розмножуватися, інші (наприклад, гепатоцити) її зберігають і після ушкодження тканини повертаються в цикл. Тривалість циклу визначається в основному довжиною періоду G1. Період G1 закінчується подвоєнням центриолей клітинного центру. Для S-періоду характерне подвоєння молекул ДНК в ядрі К. чинності неодночасність реплікації по довжині ДНК цей період триває 6-10 год. Період G2 триває 2-5 год; у цей час К. готується до митозу. У митозе (період М) відбуваються складні перетворення ядра К., забезпечують спадкоємність хромосом у ряді клітинних поколінь і поява генетично рівнозначних дочірніх клітин. У періоді Му розрізняють 4 фази (рис. 6); його тривалість 1-2 ч.
Профаза починається е розбіжності пар центриолей до полюсів клітини; між ними з мікротрубочок формується веретено поділу. Сама клітина округляється, у її ядрі конденсуються і стають видимими у світловий мікроскоп подвійні нитки хромосом. Ядерця і оболонка ядра руйнуються.
У метафазі хромосоми розташовуються в екваторіальній площині клітини у вигляді метафазной пластинки. Кожна хромосома складається з двох поздовжніх ниток (хроматид), сполучених центромерой; останні прикріплені до ниток веретена поділу спеціальними структурами - кинетохорами. Весь цей механізм, що забезпечує точний розподіл матеріалу хромосом між дочірніми клітками, називається митотическим апаратом.
В анафазі хроматиди розходяться до різних полюсам.Клетки витягується, приймає видовжену форму. З моменту поділу хроматиди отримують махання дочірніх хромосом; в кожне з дочірніх ядер потрапляє по одній хроматиде кожної хромосоми.
В телофазе фрагменти зруйнованої ядерної мембрани зв'язуються з дочірніми хромосомами; по мірі їх деконденсации ці фрагменти зливаються в загальну оболонку ядра. Відбувається реконструкція дочірніх ядер, синтез нових ядерець, руйнування мітотичного апарату. У пізній анафазі або в телофазе між дочірніми ядрами з'являється борозна ділення, або поперечна перегородка, і тіло клітини поділяється на дві дочірні К. (цитокинез цитотомия). При порушенні цитотомии мітоз може завершитися освітою дво - або багатоядерних клітин.
Особливою формою поділу клітини є мейоз, при якому двом послідовним поділом К. передує лише одноразова реплікація ДНК. В результаті утворюються статеві клітини, що містять удвічі менше ДНК і хромосом, ніж інші (соматичні) клітини організму. При злитті статевих К. (заплідненні) характерне для клітини даного організму відновлюється число хромосом (див. Розмноження, Зародок). Збільшення клітинної маси може протікати і без поділу клітини шляхом последовательною, іноді багаторазового, подвоєння молекул ДНК і, відповідно, числа хромосом у ядрі. Ці процеси називають эндорепродукцией, а які утворюються в результаті їх клітини - полиплоидными. У деяких випадках поліплоїдні або диплоїдні клітини (частіше їх ядра) поділяються без порушення цілісності ядер і ядерець шляхом прямого розподілу - амитоза. Різні форми амитоза нерідко зустрічаються при патологічних процесах (запалення, злоякісний ріст), а також в ході регенерації і старіння клітини.
Старіння клітини супроводжується порушеннями біосинтетичних процесів, включаючи транскрипцію і трансляцію генетичної інформації, а також реплікацію і репарацію ДНК. Змінюються фізико-хімічні властивості білків хроматину ядер клітини, збільшується міцність зв'язування гістонів з ДНК. Накопичення дефектних макромолекул веде до змін структури цитозолю і органоїдів і до порушення функції клітини. Різні клітини старіють по-різному. У нервових клітини, в елементах міокарда і скелетних м'язів накопичуються так звані пігменти зносу, наприклад ліпофусцин. Темп поділу короткоживучих клітини сповільнюється. Є дані, що нащадки кожної К. проходять певний, генетично детерміноване число клітинних циклів. Для фібробластів це число дорівнює 20-50. Поряд з деструктивними процесами в старіючих К. спостерігаються також пристосувальні процеси, спрямовані на відновлення функцій клітини. По мірі наростання деструкції і згасання відновлення настає природна смерть клітини.
Деструкція і смерть клітини можуть настати також внаслідок дії різних пошкоджуючих факторів. Ці процеси вивчає цитопатология. На багато видів поврежденийклетки може відповідати однотипною реакцією у вигляді комплексних змін цитоплазми (зміни в'язкості, здатності сприймати барвники), ядра (конденсація хроматину, скупчення його грудочок по периферії ядра - маргинация хроматину) і ядерець (зміна числа і розмірів, зникнення). Кількість пошкоджуючих факторів, як правило, перевищує набір можливих морфологічних змін клітини, тому визначити (діагностувати) причину пошкодження вдається лише на підставі аналізу комплексу змін сукупностей (популяції) клітини. Але іноді в реакції клітини виявляються і специфічні риси. Так, пошкодження окремих хромосом статевих клітини може лежати в основі спадкових хвороб. Структурні зміни клітини супроводжуються порушенням їх життєдіяльності. Залежно від характеру, інтенсивності і тривалості дії шкідливого агента патологічні зміни К. можуть бути оборотними чи необоротними. Виділення з ядра рідини зі стисненням хроматину у великі грудочки або в єдину масу і зі зникненням ядерця називають пикнозом. До тяжких незворотних змін ядра, зв'язаних з гибельюклетки, відносяться кариорексис (розпад на окремі грудочки) і кариолизис (вихід вмісту ядра в цитоплазму). Зміни ядра часто поєднуються з розширенням (набряком) перинуклеарного простору. В цитоплазмі спостерігаються ознаки порушення циркуляції внутрішньоклітинної рідини, зміни проникності мембранних структур клітини, гіпертрофічні та атрофічні процеси та ін. До найчастішим морфологічним проявам патології клітини відносяться набухання і руйнування мітохондрій, розпад одномембранных органоїдів на дрібні бульбашки, руйнування елементів цитоскелету. У всіх цих процесах важливу роль відіграють ферменти лізосом; у важких випадках їх мембрана розривається, і вивільняються ферменти зумовлюють руйнування (лізис) клітини.
Серед різноманітних форм клітинної патології особливо виділяють вірусну цитопатологию. Проникаючи в клітини, віруси вносять у неї чужорідну генетичну інформацію, різко змінює перебіг біосинтетичних процесів. Зокрема, вірусні иРНК зв'язуються з багатьма рибосомами клітини, формуючи вирусспецифические полисомы, на яких синтезуються білки не клітини, а вірусу. Реплікація вірусних нуклеїнових кислот і збірка вірусних частинок протікають в залежності від властивостей вірусу в цитоплазмі або ядрі клітини. Специфічними морфологічними ознаками вірусної інфекції служать вірусні включення, видимі в світловий мікроскоп. Форма, розташування, структура та хімічні особливості включень характерні для кожного сімейства вірусів, так що діагноз нерідко можна поставити на підставі вивчення окремої клітини. Ряд вірусів кору, віспи) викликає злиття клітини з утворенням багатоядерних симпластов. Комплекс змін, що передують загибелі клітини при вірусної інфекції, називають цитопатичною ефектом; віруси, що викликають загибель заражених ними клітини, називають цитопатогенными. Зміни заражених вірусами клітини лежать в основі патогенезу ряду вірусних захворювань. Специфічність морфологічних змін клітини використовують в лабораторній діагностиці вірусних інфекцій для індикації вірусу в клітинних культурах і виявлення антитіл у сироватці хворих або перенесли захворювання
Віруси, що мають липопротеидную оболонку, виходять з К. шляхом брунькування за типом екзоцитозу; при цьому клітини гине через деякий час. Інші віруси звільняються шляхом «вибуху», викликаючи швидке руйнування клітини. Вірусні інфекції, що ведуть до загибелі клітини, називають цитолитическими (цитоцидными).
Деякі сімейства вірусів здатні, не руйнуючи клітини, об'єднуватися (інтегрувати) з її хромосомами. За такої інтегративної формі інфекції вірусний геном реплікується і функціонує як складова частина геному клітини. Геном або частину геному вірусу в складі хромосом клітини. називають провирусом. Включатися в геном клітини можуть містять ДНК онкогенні віруси, аденовіруси, віруси гепатиту та герпесу. РНК-вмісні ретровіруси мають у віріона особливий фермент - зворотну транскриптазу, яка забезпечує синтез ДНК на матриці вірусної РНК. Такий ДНК-транскрипт грає роль провірусу.
Пухлинна трансформація клітини при її зараженні онковирусами обумовлена онкогенами - ділянками клітинного генома. захопленими онковирусом в ході його утворення в клітини Вірус транспортує онкоген в певне місце геному клітини, результатом чого є порушення регуляції поділу клітини.
|
 Застосування Відехолу в стоматології
Застосування Відехолу в стоматології