|
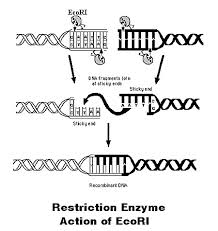
Рестриктази розпізнають певні нуклеотидні послідовності і розщеплюють молекули ДНК всередині або за межами впізнаваного ними сайту. Розщеплення призводить до дегенерації ДНК з утворенням "липких" (5`- і 3`-звисів) або "тупих" кінців.
Загальні властивості Явище специфічності дії рестриктаз була вперше виявлена Лурія і Хьюманом на початку 1950-г. Майже десять років потому, Арбер і Дюссо передбачили молекулярні основи специфічності дії рестриктаз. Вони припустили, що специфічність полягає у функціонуванні двухферментной системи: рестриктази, яка дізнається специфічні послідовності ДНК і розщеплює чужорідну ДНК при її попаданні в бактеріальну клітину і модифікуючий ферменту (метилтрансферази), який захищає ДНК хазяїна від деградації власної рестриктазою. Обидва ферменти дізнаються один і той же сайт і разом утворюють систему рестрикції-модифікації (Н-М).
Системи рестрикції-модифікації широко поширені серед бактерій, а також були ізольовані з фага, архей і вірусів эукариотических водоростей. Р-Му системи були розділені на чотири типи (I, II, III і IV) в залежності від складності їх структури, вимог кофактора, механізму дії та послідовності расщепляемого сайту. Найкраще охарактеризовані і найчастіше використовуються рестриктази II типу. Ці ферменти розпізнають певні послідовності ДНК довжиною 4-8 п. о. і розщеплюють ДНК або на сайті впізнавання, або на відстані до 20 пар підстав від сайту впізнавання.
Нерідко буває, що більш ніж один фермент дізнається певну послідовність нуклеотидів. Виходячи з номенклатури рестриктаз, фермент з унікальною специфічністю розщеплення ДНК, який був виявлений першим, називається прототипом. Згодом виявлені ферменти, що володіють тією ж специфічністю, називаються изошизомерами. Изошизомеры можуть відрізнятися від прототипу за рівнем здатності розщеплювати цільової сайт, умов реакції, а також чутливості до метилированию і спобности до зірчастої активності. Рестриктази, які визнають ту ж послідовність нуклеотидів, але розщеплюють ДНК в різних місцях називаються неошизомерами.
|
 Аварії на АЕС. Принципи обмеження радіаційного впливу на людей
Аварії на АЕС. Принципи обмеження радіаційного впливу на людей