 |
Сучасні уявлення про регуляцію менструальної функції |
Статева система жінки є саморегулюючою і включає 4 основні функції:
• менструальну,
• сексуальне,
• секреторну
• репродуктивну.
Активність репродуктивної функції, на відміну від* інших систем організму, зростає до 16-17 років, до 45 років згасає репродуктивна, а до 55 - гормональна функція репродуктивної системи. Гормональна функція організована за ієрархічним принципом, і в ній поділяють 5 ланок або рівнів, кожен з яких регулюється вищерозташованими структурами.
Менструальна функція - постійна, що циклічно повторюється підготовка організму жінки до вагітності. Вона властива лише людині і вищим приматам. Менструація - результат циклічної діяльності всієї статевої системи половозрелой.женщи-ни. Менструальний цикл - фізіологічний процес в організмі невагітної жінки, який розпочинається з першого дня останньої менструації і закінчується початком наступної, протягом якого у яєчнику відбувається дозрівання фолікула і виділення яйцеклітини, а в матці створюються умови для імплантації заплідненого яйця. Менструація - кінець менструального циклу. Середня тривалість менструального циклу 28 днів (21-35 днів), тривалість кровотечі - 1-7 днів, крововтрата - 10-80 мл
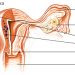
Рівні регуляції менструальної функції: вищі (кора головного мозку, гіпоталамус, гіпофіз) та нижчі рівні (яєчники, матка та інші тканини-мішені: молочні залози, піхву, волосяні фолікули, шкіра, кістки, жирова тканина). Зв'язки між вищими та нижчими рівнями можуть бути прямі і зворотні, позитивні і негативні, довгі, короткі та ультракороткі.
Вищі центри. Управління гормональною системою репродуктивної функції починається з рівня спеціалізованих нейронів головного мозку. У науковій літературі їх називають надгипоталамическими структурами. Вони отримують інформацію про стан зовнішнього середовища, перетворюють її в нейрогуморальні сигнали, які через нейротрансмиттерную зв'язок надходять у секреторні клітини гіпоталамуса. Вивчення ролі надгипоталамических структур пов'язане з певними труднощами. Вважається, що однією з них є довгастий мозок, нейрони якого виробляють норепінефріну, стимулюючий гонадотропін-рилізинг фактор (ГнРФ) гіпоталамуса. Статева дифе-ровка починається з гіпоталамуса. Він є частиною проміжного мозку. Так звана гіпофізарна зона гіпоталамуса розташовується на підставі третього шлуночка мозку (вентро - і дорсомедиальные ядра). В них утворюються либерины, за хімічною структурою належать до декапептидам.
Гіпоталамус - центральна ендокринна залоза. Відомі шість тропных гормонів: стимулюючі - либерины, гальмують - статини. В даний час існує концепція про єдиний либерине, який є стимулюючим гормоном для лютеонизирующе-го гормону (ЛГ) та фолікулостимулюючого гормону (ФСГ) і називається люлиберином. Він надходить у кров у пульсуючому цирхоральном режимі: 1 пульсація за 60-90 хвилин. У залежності від частоти і амплітуди змінюється концентрація ЛГ і ФСГ. Кожен секреторний імпульс люліберіну не пізніше, ніж через 2-5 хвилин супроводжується підйомом рівня ЛГ в крові. Якщо пульсація люліберіну ГнРФ збільшується в 2 рази, то вміст ЛГ знижується в 2 рази, а ФСГ - на 65%. При подальшому збільшенні частоти пульсації люліберіну (3-5 разів на годину) кількість ЛГ і ФСГ різко знижується, відсутні їх пульсуюча секреція. Це пояснюється перенасиченням рецепторів гонадотро-фов. При цирхоральном ритмі пульсації зв'язує здатність встигає відновитися. Таким чином, люлиберин є либерином і статинів для двох гонадотропних гормонів.
Для пролактину либерином є тиреотропин-рилізинг-гормон, статином - пролактин інгібуючий фактор (ПІФ) - дофамін (ТАК). Однак, либерин має деякий вплив на пролактин. Цирхоральный ритм секреції люліберіну призводить до цирхоральному ритму секреції гонадотропінів, але абсолютна співвідношення цих гормонів у різні дні менструального циклу впливають гонадальные гормони. Зв'язку між надгипоталамическими структурами гіпоталамусом і гонадальными гормонами здійснюється через нейротрансмітери. До них відносять норадреналін (НА), серотоніну, ТАК, ЕОП (ендорфіни, энкефа-ліни). НА стимулює викид люліберіну, серотонін - гальмує його. ТАК - стимулює люлиберин і пригнічує секрецію пролактину.
Ендогенні опіоїдні пептиди (ЕОП) утворюються внаслідок розщеплення молекул-предщественниц, що дають початок відповідно трьом групам: ендорфіни (ЕНД), енкефаліни (ЭНК), динорфінів (ДІН). ЕОП є не тільки в межах мозкової тканини, але і в інших органах і тканинах. В яєчниках, наприклад, концентрація ЕНД значно вище в жовтому тілі, що дозволяє припустити їх роль у функції жовтого тіла, у фолікулах їх менше, а в примордиальных фолікулах відсутні зовсім. Більше всього вивчено їх вплив на гонадотропну функцію гіпофіза. ЕОП гальмує секрецію люліберіну.
Через ЕОП відбувається вплив гонадальных гормонів на люлиберин і, як наслідок, на ЛГ і ФСГ. (3-ендорфін в свою чергу призводить до стимуляції секреції пролактину. Дослідженнями доведено, що ЕОП впливають на пролактин, знижуючи секрецію ТАК. Серотонін стимулює ПрЛ і знижує концентрацію люліберіну.
Простагландини - особливі внутрішньоклітинні речовини, похідні поліненасичених жирних кислот, звільняються і діють в клітці 20 видів. Їх попередником є арахідонова кислота. По мірі насичення вони діляться на простагландини, простациклины, тромбоксаны. Останні два - метаболіти простагландинів. Вони змінюють концентрацію циклічного аденозинмонофосфорной кислоти та іонів кальцію. Естрогени стимулюють вироблення простагландинів, прогестерон і пролактин пригнічують їх.
Наступним, третім рівнем регуляції є гіпофіз. Аденогіпофіз виробляє ЛГ, ФСГ, ПрЛ, регулює функцію яєчників і молочних залоз. ЛГ та ФСГ - гликопротеиды, ПрЛ - поліпептид. Гіпоталамус і аденогіпофіз знаходяться в тісній функціональній залежності. ФСГ стимулює ріст фолікула, проліферацію гранулезных клітин, продукує утворення рецепторів до ЛГ на поверхні клітин гра-нулезы. Під його впливом дозріває гранулеза, стимулюється ароматаза, синтезуються естрогени. Однак для виділення естрогенів необхідний ЛГ. Тільки при спільному впливі ФСГ і ЛГ відбувається дозрівання фолікула. Піковий викид ЛГ відбувається при високій концентрації естрадіолу в крові. ЛГ надалі стимулює функцію жовтого тіла і разом з пролактином сприяє секреції в гранулезо-лютеиновых клітинах прогестерону. ЛГ стимулює тека-тканина яєчників, які секретують андрогени (андростендіон, тестостерон).
Основна роль пролактину полягає в стимуляції розвитку молочних залоз, лактації. Крім того, він володіє жиромобилизующим ефектом, під його впливом відбувається стероїдогенез. Оптимальний рівень пролактину сприяє стероидогенезу, тоді як підвищений або знижений рівень гальмує його.
Для гіпофіза характерні 3 режими секреції (тонічний, цирхоральный, циклічний). Пульсуюча секреція люліберіну необхідна для індукції ЛГ і ФСГ, тоді як рівень гормонів в крові, їх оптимальне співвідношення підтримується процесом, що відбувається на рівні гіпофізарно-яєчникових взаємозв'язків. Таким чином, пульсуюча секреція люліберіну підтримує постійний цирхоральный ритм секреції гонадотропінів (ГТ). Зміна рівня ГТ і їх характеру секреції протягом циклу залежить від гонадальных гормонів. Прогестерон знижує частоту викидів ЛГ, збільшуючи їх амплітуду, естрадіол ж знижує амплітуду, не впливаючи на частоту. Естрогени діють на гіпофіз, прогестерон впливає безпосередньо на гіпоталамус (аркуатные ядра).
Другий рівень регуляції репродуктивної системи - яєчники. У них відбувається розвиток фолікулів і стероїдогенез. Процес фолликулогенеза відбувається постійно, починається антенатально і закінчується постменопаузально. Піддаються атрезії 90% фолікулів і лише 10% проходить повний цикл: дозрівання, овуляція, жовте тіло. Фолікули від примордиального розміром 2 мм ростуть до 10 мм під впливом пара-кринных факторів, подальший фолликулогенез здійснюється під впливом ФСГ. Під впливом ФСГ відбувається розвиток домінантного фолікула, який до 14 дня досягає 18-20 мм в діаметрі. Фолікулярна рідина збільшується стократно. Гранулеза починає ділитися: з одного шару клітин, що вистилає soupe фолікул, перетворюється в потужну гранулезную мембрану. Фолікули, стінки яких складаються більш ніж з одного шару клітин, називаються вторинними.
Знаходиться в порожнині ооцит ооцитом називають першого порядку. Він зростає, збільшуючи свій розмір, навколо нього розвивається мембрана - Zona pellucida (блискуча оболонка). Утворюється фолікулярна рідина, яка відтісняє фолікулярні клітини і ооцит до однієї сторони, утворюючи яйценосний горбок. У фолікулярній рідині різко збільшується концентрація естрадіолу і ФСГ, накопичується інгібін Ст.
В організмі жінки існує 3 основних естрогену: естрон, естрадіол, естріол. Естрон і естрадіол первинно утворюються в яєчниках. Вони можуть взаємно перетворюватися один в одного і можуть перетворюватися в естріол. Але останній не може перетворюватися в естрадіол і естрон, а виводиться з сечею. Естріол не має проліферативним ефектом і малоактивний. Максимальний рівень естрогенів визначається перед овуляцією. Другий їх пік припадає на час фази розквіту жовтого тіла. Попередниками в їх синтезі є андрогени: андростендіон є попередником естрадіолу, тестостерону - естрону. Специфічна здатність естрогенів полягає в стимуляції проліферативних процесів в ендометрії. Вони покращують кровообіг і трофіку матки, підвищують чутливість рецепторів до окситоци-ну, впливають на розвиток вторинних статевих ознак, сприяють плісняві епіфізів трубчастих кісток. Естрогени (B-естрадіол) через інгібін У пригнічують секрецію ФСГ і зумовлюють овуляторний викид ЛГ. Отже, після завершення росту фолікула й согревательного поділу яйцеклітини (ооцит другого порядку) відбувається овуляція, яка і завершує першу фазу менструального циклу. Овуляція відбувається під впливом колагенази, простагландинів F2 і Е2, протеолітичних ферментів, окситоцину, релаксину. В фолікулярної рідини перед овуляцією визначається значне зміст ингибина - речовини, що пригнічує секрецію ФСГ.
На процес овуляції вирішальний вплив мають: рівень 17 B-естрадіолу, піковий викид ЛГ, простагландини. Для овуляції необхідна синхронізація двох процесів: високий рівень естрадіолу і піковий викид ЛГ. Якщо має місце більш раннє дозрівання фолікула (або запізнювання викиду ЛГ), настає персистенція фолікула. Запізніле дозрівання фолікула (або передчасний викид ЛГ) призводить до атрезії фолікула. Час овуляції постійно і становить 14-16 днів до початку нової менструації.
Після овуляції настає друга фаза - лютейновая. Після того, як ооцит другого порядку і деяку кількість рідини изверглись при овуляції фолікул спадается. У ньому відбувається невелике крововилив з утворенням невеликого згустку. Залишилися фолікулярні клітини збільшуються в розмірах і перетворюються в фолликулярно-лютеиновые клітини. Жовтий пігмент, накопичуючись у них, надає їм характерний жовтий колір. З цього моменту жовте тіло набуває функцію ендокринної залози і продукує прогестерон під впливом Л Р, ПрЛ.
Жовте тіло секретує прогестерон 12-14 днів, після чого, якщо не настала вагітність, починається його інволюція. Прогестерон веде до секреторної трансформації ендометрія, до децидуальної реакції строми. Головна його функція полягає у підготовці матки до імплантації заплідненого яйця. Відзначено також діуретичну і термогенное вплив прогестерону через гіпоталамус - температура тіла піднімається на 0,6-0,8°С. Гормон здатний гальмувати ЛГ, впливаючи на гіпоталамус.
I рівень регуляції - матка. Ендометрій матки в кожному циклі проходить кілька фаз:
• з 1 по 4 день - менструальна фаза,
• з 4 по 12 день - проліферативна (фолікулярна) фаза.
Ендометрій збільшується з 1 до 5 мм. Зростання його стимулюється естрогенами (функціональний шар). З 13 дня під впливом прогестерону настає фаза секреції. Таким чином, дві фази яєчникового циклу відповідає двом фазам маточного. Фаза дозрівання фолікула - фазі проліферації, фаза жовтого тіла - фазі секреції. Отже, нормальний менструальний цикл - це двофазний цикл.
До першого рівня регуляції, крім матки, як вже згадувалося, відносять молочні залози, волосяні фолікули, шкіра, кістки, адипоцити. Клітини цих тканин містять рецептори до статевих гормонів.
Цитозолрецепторы - рецептори цитоплазми, які мають суворої специфічністю до эстрадиолу, прогестерону, тестостерону. Вільна молекула стероїдного гормону захоплюється специфічним цитозолре-цептором білкової природи і що утворюється комплекс транслоцируется в ядро клітини. В ядрі виникає новий комплекс з ядерним рецептором. Цей комплекс зв'язується з хроматином, що регулює транскрипцію мРНК, які беруть участь у синтезі специфічного тканинного білка.
До першого рівня репродуктивної системи відноситься внутрішньоклітинний медіатор циклічна аденозинмонофосфорная кислота - (цАМФ), яка регулює метаболізм в клітинах тканини-мішені у відповідності з потребою організму у відповідь на дію гормонів. До першого рівня відносять і міжклітинні регулятори - простагландини. Їх дія реалізується через цАМФ.
Відомо вплив надниркових залоз щитовидної залози, епіфізу, печінки на репродуктивну систему, тому при появі дисбалансу у функції будь-залози внутрішньої секреції відбувається поломка в гормональної функції репродуктивної системи.
Таким чином, ендокринна система жінки - це єдина система і лише при оптимальній її функції залоз досягається нормальний гормональний гомеостаз.
|
 Чудодійний корінь імбиру
Чудодійний корінь імбиру

