Меланосоми (від грец. melas, рід. падіж melanos - чорний і сома), цитоплазматичні структури меланоцитів і меланофорів, на білковому матриксі яких синтезуються пігменти меланіни і відкладаються у вигляді крейди-протеїнових комплексів. У розвитку меланосом розрізняють декілька стадій премеланосом різних ступенів зрілості (по заповненню меланіном). Повністю заповнені меланіном меланосоми, видимі в світловому мікроскопі, називаються меланіновими гранулами.
Мал. 1 Мал. 2
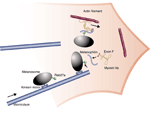
Еукаріотичні клітини у великій кількості експресують моторні білки, які беруть участь у Широм колі клітинних процесів. Kinesins, dyneins і myosins використовують енергію, що вивільняється при гідролізі АТФ (ATP), щоб переміщати свій вантаж уздовж мережі цітосклета, актівнових мікрофіламентів і мікротрубочок. Связиваеніе забезпечується декількома класами адапторною 'рецепторних' молекул. Одним із класів є Ras-подібні GTPases сімейства Rab. Майже 60 членів сімейства Rab ідентифіковано в клітинах ссавців. Шляхом взаємодії з широкою різноманітністю ефекторів, вони відіграють ключову регуляторну функцію по координації последовательнчих стадій мембранного транспорту, включаючи формування транспортних носіїв і їх зв'язування / закріплення з мембранами-мішенями. Зв'язок між Rab і моторними білками вперше продемонстрована на Rab6a, асоційоване з Golgi Rab, який залучений в ретроградний транспорт між Гольджі і ендоплазматичним ретікулемом. Rab6 взаємодіє з kinesin-подібним білком Rabkinesin-6. Показано, що та ін Rab білки, включаючи Rab5 і Rab11, м. взаємодіяти безпосередньо з базуються на мікротрубочки і актине моторами. Wu та ін детально описують як Rab білок (Rab27a) використовується моторним білком (myosin-Va) в органелах (Меланосоми) Меланосоми формуються в меланоцитах, локалізованих в підставі волосяної цибулини і в базальному шарі епідермісу. Біогенезу меланосом включає багато подій сортування на endocytic і секреторних шляхах. Зрілі меланосоми переміщаються уздовж мікротрубочок від околоядерном Гольджі області в напрямку периферії клітин (дендритів) шляхом взаємодії зі звичайним кінезіном (kinesin II у Xenopus laevis). Досягнувши периферії, меланосоми від'єднуються від мікротрубочок і зв'язуються з субкортікальной Актинові пучками через взаємодію з міозином Va. Меланосоми і їх вміст, меланіновий пігмент, передаються оточуючим кератиноцитам. Цей процес важливий для пігментації шкіри і волосся. Критична функція міозину Va у підтримці меланосоа вперше була виявлена у мишей, коли було встановлено, що ген забарвлення шерсті dilute кодує важкий ланцюг міозину Va. У меланоцитах, виділених від мишей, гомозиготних по нульовому аллелю dilute, меланосоми накопичуються в околоядерном зоні і не передаються кератиноцитам. Це пояснює дефект пігменту у dilute мишей. Нещодавно було встановлено, що синдром Griscelli, рідкісне аутосомно імунне захворювання асоційоване з частковим альбінізму волосся і шкіри, виникає в результаті мутації в гені myosin-Va. Отже, синдром Griscelli у людини еквівалентний dilute у мишей. Далі було встановлено, що у ряду пацієнтів з Griscelli розвивається неконтрольований синдром активації T-lymphocyte і макрофагів, що мають крім того мутації в гені RAB27A. В той же час було повідомлено, що мутантний ген у мишей ashen, який, як і у випадку dilute, викликає дефекти пігментації через аномальне транспорту меланосом, є rab27a. Потім було встановлено, що разом з міозином Va, Rab27a присутній на поверхні меланосом і що порушення його функції викликають у мишей і людини порушення розподілу меланосом Відсутній шматок мозаїки знайдений, це melanophilin, продукт гена leaden. Leaden миші виявляють порушення забарвлення шерсті, відрізнити від таких у ashen або dilute. Встановлено, що melanophilin асоціює зі зрілими меланосоми, локалізованими на периферії мелоноцітов. Як і очікувалося, melanophilin-позитивні меланосоми мітяться також і по Rab27a і myosin-Va. Leaden меланоцити накопичують меланосоми в околоядерном області клітин, цей фенотип виявилося можливим нормалізувати тимчасової трансфекції melanophilin. Встановлена ієрархія подій, що відповідають за правильний таргеттінг Rab27/melanophilin/myosin Va в меланосоми і їх myosin Va-залежне відлов на периферії клітини: по-перше, екзогенний міозин Va не направляється в меланосоми в leaden меланоцитах, по-друге, міозин Va не виявляється на поверхні меланосом в ashen (rab27a-) клітинах, трансфіцірованних melanophilin; по-третє, Rab27a направляється в меланосоми як dilute нульові меланоцити, так і в leaden меланоцити. Все це вказує на те, що спочатку Rab27a з'єднується з меланосоми, потім melanophilin і, нарешті, міозин Va. Щоб виявити молекулярні зв'язки між Rab27a, melanophilin і myosin Va, авт. аналізували патерн взаємодій різних конструкцій. Взаємодія Rab27a з melanophilin є GTP-залежним і використовує N-термінальний домен (амінокислоти 1-152) melanophilin. Цей домен містить 8 законсервованих цистеїнових залишків, які утворюють домен цинкові пальчики і мотив six-aromatic-amino-acid-rich, виявляється у різних Rab ефекторів, закінчує Rab зв'язує домен. Домен, що взаємодіє з myosin Va, у melanophilin розташовується на його С-кінці. Цікаво, що ця взаємодія потребує присутності Exon F, альтернативно сплайсіруемого екзонів, кодує 25 амінокислот в хвості міозину Va. Exon F присутня в меланоцит-сплайсірованной ізоформи міозину Va, але не в ізоформи, сплайсіруемой в головному мозку. Цього, однак, недостатньо для взаємодії melanophilin-myosin Va, тому це взаємодія потребує також в участі глобулярного хвоста міозину Va. Викликали надлишкову експресію або N-або C-термінальної частини melanophilin в меланоцитах дикого типу. Відповідно до передбачуваної послідовністю подій (Мал. 2), обидві конструкції витісняли міозин Va з меланоцитів і створювали leaden-подібний фенотип. Цікаво, що Rab27a зв'язується на Меланосоми епітеліальних клітин пігментного епітелію сітківки з ін actin-based мотором, міозином VIIa, за допомогою білка, званого MyRIP (myosin VIIa and Rab interacting protein), який схожий з melanophilin. Подібним чином, Rab11a ефектор, Rab11-family-interacting protein 2 (FIP2), взаємодіє з міозінових Vb хвостом, а Rab11-Rab11 FIP комплекс служить як рецептора для міозину Vb на поверхні реціклірующіх ендосом. Можливо, що будуть виявлені й ін приклади Rab-моторний білок взаємодій. Ідентифікація множинних Rab ефекторів з широким спектром біологічних активностей ілюструє чудовий функціональний репертуар Rab GTPases. Передбачається. що Rab ефектори прдеопределяют і організують домени на поверхні внутрішньоклітинних компартментов, роблячи їх компетентними для функцій, таких як злиття вхідними транспортними проміжними структурами або ін органелами, а також для ремоделювання органел. Фактично, функція Rab GTPases скоординована як просторово, так і в часі з процесом утворення доменів мембран, із взаємодією і / або рухом цих домено уздовж цитоскелету. Rab GTPases м. розглядати як центральні регулятори динаміки органел у клітинах еукаріот.
|